The Emergence of Disease
in Early World-Systems:
A Theoretical Model of
World-System and Pathogen Evolution[i]
Anthony Roberts
Department of Sociology and
Institute for Research on World-Systems
(IROWS)
For presentation at the annual meetings of the
American Sociological Association, Atlanta, Georgia, August 15, 2010, PEWS
Roundtable on the Evolution of World-Systems. This is IROWS Working Paper #62 available at https://irows.ucr.edu/papers/irows62/irows62.htm
Draft v. 8/8/2010 Words=5,740
Abstract
In
theorizing socio-cultural evolution, through demographic-based perspectives, a
central component regulating human populations is often absent. The model offered in this theoretical study
introduces the component of disease emergence and re-emergence as a central
regulator of human populations in an early horticultural society embedded in
regional world-systems with hunter-gather societies. Based on a formalized version of the Human
Demographic Regulator model (Apkarian et al 2010), I incorporated an SIR
disease modular into the local population system to explore how disease
regulates human populations to sustainable levels for the local resources and
how disease, as a regulator, interacts with inter-societal processes,
emigration and warfare, to regulate human populations. A formalized version of the modified HDR
model was simulated based on different pathogen characteristics (transmission
risk and mortality) to uncover population and disease dynamics. According to the model, societies with
moderate domestication rates generate the appropriate conditions for a pathogen
with high virulence to sustain itself.
Further, disease operates in tandem with other regulators identified in
the original HDR model, however, if the pathogen is deadly and easily
transmitted, than disease becomes the primary regulator of human
populations.
In developing explanations of disease emergence,
re-emergence, and spread, insufficient attention has been paid to long-term
socio-historic processes as primary articulators of disease dynamics. To compensate for this deficiency in
epidemiology and other disciplines, this article seeks to incorporate basic
disease dynamics in macro-structural modeling of early human societies, in
particular, societies transitioning from simple sedentary societies based on
diversified foraging modes of production to complex sedentary societies based
on basic horticultural modes of production.
By modeling disease dynamics in simple society evolution, in roads are
developed to understand how the intensification of social relations, the
reduction of spatiality, and the formation of global institutions impact
disease dynamics. In a precarious
period of human health, a better understanding of social systems and disease
could shed light on the direction of human health in a globalizing world.
For understanding the relationship
between the environment and human populations, a systemic approach is necessary
to account for the mutual dependency and articulation between social and
ecological systems. Social systems, as systems embedded in ecological systems,
must adjust to the conditional parameters of the environment (Williams 2003;
McNeil 1976; Odom 1975). According to a
Malthusian hypothesis of exponential population growth and arithmetic resource
growth, there is a real ecological limit to size of human populations, thus,
demography is a critical factor in the environment-human nexus. However, over the long history of human
society, social systems have increasingly pushed the boundaries of ecological
constraint through expanding populations and resource intensification resulting
in depletion, pollution, and other negative externalities.
One of the answers on how social systems
regulate human populations is the Iteration model (Chase-Dunn and Hall 1997, P.
102) and its formalized version, the Human Demographic Regulator (HDR) (Apkarian
et al 2010). According to the HDR model,
human populations are regulated by resource limitations, warfare, and
migration. Absent in the theory,
however, is the role of epidemics, which came about with the transition from simple
horticulture and foraging to complex horticulture in early human societies. This article proposes a formalized model
extending from the HDR model by introducing disease dynamics as a regulator of demographic
pressures on the environment.
Using formalized
theory modeling, three variables are isolated as primary mechanism in the
human-disease relationship in the HDR-disease model presented in this paper:
transmission risk, mortality, and domestication (human-animal relations). The purpose of formalizing the modified
iteration model (HDR)[1] is to
conceptualize how diseases became a primary regulator in early, simple
sedentary societies and how disease dynamics relate to inter-societal relations
between a local social system and a regional inter-societal system.
Theoretical finding from the model indicates
in social systems with moderate to high domestication, the primary regulator is
the emergence and re-emergence of crowd disease[2]. Under certain conditions, specifically the
presence of a developed regional system, warfare operates as a secondary
regulator in control the demographic growth of local populations. Further, a counter-cyclical trend emerges
between warfare and disease, where warfare operates as a suppressant of disease
reproduction in the local population.
Migration, in the modified model, is
primarily driven by the rate of domestication and the virulence of the
pathogens. In resource rich scenarios
with high domestication rates, population densities grow substantially,
resulting in frequent waves of migration.
Under conditions with high disease lethality, however, migration never
occurs because population pressures are minimized. Given the theoretical results of the
formalized model under different disease conditions, the tentative conclusion
of the study is, with the emergence of domestication in sedentary societies, an
intensification of the interaction of social and biological systems occurred,
which results in a co-dependence between pathogens and humans. Additionally, the synchrony of warfare and
disease provides some theoretical validity to the relationship between
world-systems and biological systems.
Disease and Domestication
A central focus of the modified HDR theory is changes in
human populations caused by the biological operation of pathogens and
inter-societal relations in a small world-system composed of a sedentary
society, with increasing population density, and a regional system of
societies. The original version of HDR
developed by Apkarian et al (2010), concentrates on the relationship between
human populations and local eco-systems.
In the formalized model, resource production and consumption are the
core drivers of demographic dynamics in both local and regional populations. The primary factor identified in the single
society demographic regulator (SSDR), for accounting for population variation
and dynamics, is consumption per capita, which is a function of both resource
production and population density. If
human populations exceed the carrying capacity of the local resource system,
where consumption is exceeding reproduction, than internal conflict,
starvation, and other endogenous factors reduce populations to sustainable
levels. The introduction of an
interacting regional system to the SSDR (WSDR) creates several more factors for
regulation demographic pressure, specifically, emigration and warfare. A mediating function for both
inter-societal factors (warfare and migration) is the degree of
circumscription, or the barriers to movement (Apkarian et al 2010).
The purpose of the modified theory is to
introduce a new factor in demographic dynamics, which arose out of the
intensification of resource exploitation (domestication) associated with the
first stage of human socio-cultural evolution – the emergence of crowd disease
in sedentary societies (McNeil 1976, Ch. 2; Diamond 1997, 2002; Wolfe, Dunavan,
Diamond 2007). Besides the factors
specified in the HDR model for explaining changes in populations in simple
societies, the prevalence of disease is one of the main causes of morbidity and
mortality in early human societies[3] (Dobson
and Carper 1996). By introducing disease into the demographic
systems of the local population, the advent of domestication and the emergence
of crowd disease can be theorized alongside other factors previously specified
in the original HDR model. Through
modeling disease in socio-cultural evolution, form a demographic perspective,
we can better understand how biological systems and human social systems
operate conterminously.
The origin of the intensifying
relationship between human populations and the environment is marked by several
significant ruptures resulting in a transformation of the relationship. In simple societies, the significant rupture
was the transition from hunting and forging toward domestication and complex
agriculture cultivation. The rise of
animal domestication and plant cultivation is associated with parallel social
and demographic changes, specifically, the development of advanced
technologies, expansion of human populations, and changes in ecological
conditions (Cohen 1977; Flannery 1969; Diamond 1997). Domestication of plant and animal life was
an adaptive innovation of human social organization to address environmental
pressures imposed by exogenous and endogenous processes from both social and
ecological systems.
Prior to domestication and sedentism in
human social system, the primary mode of organization and production was
nomadic hunter-gathering. In context
of infection dynamics, the low population density of hunter-gathering societies
was not conducive to the reproduction of disease at endemic levels (McNeil
1976, P. 53). For disease to adapt to a
host population, the rate of transmission is critical. Low-frequency contact, due to low population
densities, in host populations reduces the likelihood of transmission of
virulent pathogens (Ibid, P. 21).
Further, in addition to human-human contact, nomadic hunter-gatherers
had low-frequency contact with animal host populations. Therefore, while disease existed in
hunter-gathering society, the lack of transmission reduced the adaptability of
pathogens to sustain itself in human host populations. Rather large social animal populations
served as better hosts for pathogens because they had the requisite population
density needed for effective transmission and pathogen reproduction (Diamond
1997). The change from nomadic
hunter-gathering to sedentary domestication was the necessary change in human
socio-cultural organization for pathogens to adapt to humans as host populations.
The domestication of natural resources
has been associated with a variety of factors from changes in climate to
changes in population density. Gupta
(2004) proposes the emergence of domestication is associated with a change in
resource variability caused by climate amelioration in the early Holocene. Favorable climate conditions, according to
Gupta (2004), explains the geographic concentration of early domestication,
where changes in climate increased the availability of diversified and growth
of wild plants, which aided early horticulturalist in domesticating wild
plants. Diamond (1997), argues that
the emergence of domestication is based on the decline of resource
availability; resource depletion; technological change; and an increase in
population density. The initial cause
of domestication is still under debate, but all explanations converge toward
theories oriented around the human-environment relationship, rather than
exogenous forces and/or moments of human innovation. The advent of domestication was propelled by
the change in the resource-human nexus.
Due to the lack of an approximate mechanism for the rise of
domestication, the modified HDR model includes domestication as an exogenous
variable (no other variable causes change in domestication).
In human evolution, domestication is one
of the primary epochal changes in social organization and the human
evolution. By 6000 B.C., the
domestication of cattle, sheep, goats, pigs, and dogs was achieved in certain
human populations (Crabtree 1993).
Prior to the full-scale sedentism, a hybrid of horticultural-forging
modes were common, thus, full domestication was not achieved until later in
human evolution. The intensification of
animal domestication occurred 10,000 to 7,000 ago (calibrated BP years) (Gupta
2004). For example, the domestication
of cattle occurred about 7,800 years ago in present-day
Insight from medical anthropology
contextualizes the implications of domestication in the evolutionary processes
of pathogens by interpreting changes in demographic and social changes as
primary drivers of disease emergence and sustainability (Armelagos and Barnes
1999; Woolhouse and
Gowtage-Sequeria 2005; Woolhouse,
Haydon, and Antia 2005). The epidemic
transition associated with domestication (Armelagos, Goodman, and Jacobs 1991),
created a paradoxical situation for human populations – the transition from
forging and hunting modes of production to complex cultivation and animal
domestication, which produced a surplus of resources to support higher human
populations and greater population density, created a major transition in
pathogen evolution, which caused a substantial decrease in human health. A growth
in human population density created the necessary conditions for long-term
survival of disease.
However, the change in human population structures
is not the only condition for the emergence of early endemic crowd diseases in
human societies. McMichael (2004)
argues that the central cause for the emergence of zoonotic pathogens was the
growth in animal population density accompanied with domestications. As primary vectors, domesticated animals and
mammalian-dependent animals (e.g. mosquitoes, rats, etc.), especially social
animals (Diamond 1997), were highly effective transmitters of many of the
zoontic diseases found in early sedentary societies (Greger 2007). A central component in accounting for the
emergence of crowd disease is the significant intensification of human-animal
contact. The domestication of wild life
increased the frequency of human-animal interaction and facilitated the
environmental conditions suitable for disease reproduction in animal
populations (limited space, contact with waste, and several other
factors).
A modern case of pathogen and social
system co-evolution is the malaria virus.
Hume, Lyon, and Day (2003) found the introduction of domestication and
resulting increases in population density created the necessary conditions to
sustain plasmodium falciparum in human populations. Further, through human vectors, the spread
of the parasite was primarily driven by successive waves of human migration
into regional spaces. Especially for
vector-dependent transmission pathogens, like Malaria, the rise of animal
domestication and plant cultivation is one of the primary factors in the emergence
in parasitic and zoonotic disease (c.f. de Zulueta 1994). Malaria provides one of the best
illustrations of the dependency of human evolution and pathogen evolution began
by the rise in contact between animal vectors and dense human populations.
Similar to the human-environment
relationship, the survivability pathogen-human relationship is dependent on the
ability of the pathogen to sustain its environment while transforming the
environment for its survival. In the
evolution of disease strain, the balance of virulence and transmission is a
primary dynamic in the sustainability and fitness of pathogens. However, beyond the basic dynamic, there is
much debate over the exact relationship between host sustainability and
virulence (c.f. Alizon, Hurford, and Baalen 2009). In general, pathogens with a high virulence
are unable to reproduce at sustainable levels in the long-term because of high
mortality, thus, diminishing susceptible populations for transmission. Low mortality indicates a smaller degree of
host exploitation and the risk of avirulence through immunization in host populations. Based on the reproduction fitness of
disease, the emergence of crowd disease could only be possible in with changes
in human population structures, where human reproduction and immunization
created endemic conditions. This
explains the lack of crowd disease in hunter-gathering societies, where
population density was too low to sustain virulence over the long-term.
In general, the purpose of the article is to
merge demographic-based theories of socio-cultural evolution, specifically, the
HDR model, with disease dynamics. The transition
from hunter-gathering societies to simple horticultural societies represents a
novel moment in the evolution of human societies, one marked by a central
pathway in the human-environment relationship – the emergence of endemic
disease. By grafting theories of
epidemic population dynamics onto the HDR model, the association between
demographic dynamics, caused by disease, and social dynamics can be better
understood. More specifically, and the
novel contribution of the disease-HDR model, is the formalization of the
relationship between disease population dynamics and inter-societal
relationships. Based on the mechanisms
and assumptions of the HDR, if early inter-societal relations were driven by
demographic pressures and resources, than, disease should have a direct impact
on the development of inter-societal relations.
Disease as a Demographic
Regulator
In simple sedentary societies, the degree of domestication
is one of the primary factors in resource production and demographic pressures,
in conjunction with natural climate and environment. The evolutionary process of domestication
signified an upward shift in the production frontier of simple sedentary
societies by providing limited control over the reproduction resources in the
local environment. Each upward shift, with greater technology and ability to
domesticate/cultivate resources, created a stepwise development of food
production (Diamond 1997). The greater
yields gained by the domesticated plants and animals facilitated the rapid
population expansion in human societies, facilitating the conditions for
urbanization, social complexity, and other socio-cultural changes with each new
upward shift in the production frontier (Ibid). Over the course of human social
development, especially in early stages of simple societies, the degree of
resource production and consumption are principle directors of demographic
dynamics (Apkarian et al 2010). Change
in resource production, which creates a growth in consumption, expands the
carrying capacity of local environments, allowing for larger populations.
As
discussed earlier, the main model utilized in the theoretical study of disease
dynamics and socio-cultural evolution is a formalized, mathematical model of
the Human Demographic Regulator (Apkarian et al 2010; Chase-Dunn and Hall
1997). The model is divided into two
patches: a local society patch and a regional ‘societies’ patch. Both patches are designed as modified Lotka–Volterra
predator-prey models, where human populations prey on resources. Based on Chase-Dunn and Halls’ (1997)
iteration model, the HDR is a non-linear dynamic system for endogenous
processes (regional population-resource systems; local population-resource
systems) and exogenous processes (regional-local warfare, local-regional
emigration).
Temporal dynamics of the model are
incorporated in the iterative framework, where the endogenous and exogenous
processes are repeated over five thousand iterations. The central relationships specified by the
HDR model are the ‘checks’ on population pressure, specifically, internal
warfare, ‘external’ warfare, and emigration.
As the local society’s population approaches carrying capacities, the
system implements “checks” on the growing population to maintain an equilibrium
between local resource capacity and populations. The modified model offered in this study
introduces a new endogenous check on population – disease mortality.
The main
modification to the HDR model is the application of an SIR model (Susceptible-Infectious-Recovery)
to the local society patch. The SIR
model is a conventional formalization based on compartmentalizing human
populations into three vectors at different stages of disease development. Based on the differential equations developed
in an earlier mathematical epidemiology study (Kermack and McKendric 1927), the
key dynamic is the pathogen reproduction rate (Ro), or the number of
people a single infected person will infect from the susceptible population. If
R0 <1, than the disease will die out; if R0 > 1
than the disease is an epidemic; and if R0 = 1, than the disease is
endemic in the host population. Equation
1 is the rate of infection used in the model:
(1) ![]()
Where,
I is total infectious population, S is the total susceptible population, and N
is the total local population. β
represents the effective contact rate formalized in Equation 2:
(2) ![]()
Where
the effect contact rate is a multiplicative function of ![]() , the transmission risk, and contact
rate, where p is local human population density and a is local animal human
population density.
, the transmission risk, and contact
rate, where p is local human population density and a is local animal human
population density.
Besides
the relative size of the infectious and susceptible population, the key factor
in disease reproduction is the effective contact rate. In the SIR model, effective contact is composed
of two components: contact rate and transmission risk. Contact rate is a function of human
population density and animal population density, where the total contact rate
is the summation of the probability of human-human contact and animal-human
contact rate.
Transmission risk is an exogenous factor,
which varies from .02 to and 1, where .02 is a two percent chance of
transmission and 1 is a 100 percent chance of transmission. The purpose of leaving transmission risk an
exogenous factor is to retain the capability to adjust the model to specific
pathogen strains. Transmission rates
vary depending on the nature of transmissions.
For example, frequency dependent diseases, such as STDs, vary in
transmission risk depending on the behavior of the social actor (Boots and
Sasaki 2003).
An important assumption in the SIR
modular is that the recovered population gains immunity to the disease through
contraction and recovery. Another assumption
of model is there is no cross-generation immunization, where immune parents
pass on immunity to children, rather, the only way to gain immunity is to be
exposed and infected with the disease.
Immunization plays a critical role in human development. Further, the model does not incorporate any
genetic adaptation to the disease.
Immunity to deadly pathogens has played a central role in the expansion
of European populations across the globe (Diamond 1997), thus, representing a
key component in human socio-cultural evolution. However, due to the extreme
complexity associated with immunization and other genetic adaptation to
diseases, the model does not account for genetic changes in the local
population.
Disease
mortality in the model is an exogenous factor and a probabilistic function of
consumption per capita. High
consumption is assumed to translate into healthier populations, which causes an
increase of 20% in the rate of recovery from the disease. Normal recovery rate, or virulence, is an
exogenous variable ranging from 0% to 100%.
The purpose of not endogenizing virulence is to all for tests of
different strains with greater or less mortality.
In addition
to the SIR modular, the modified HDR model incorporates domestication rate as
an exogenous factor influencing animal population density, resource
reproduction, and harvesting rate. The
purpose is to model the upward shift in resource production due to the rate of
resources domesticated by the local population. Due to the ambiguity of the casual
processes for domestication, the variable is left exogenous to model. Indirectly, domestication relates to disease
dynamics through increasing the contact rate (increasing animal population
density and human population density). Human
population densities increase with greater resource production and increasing
birth rates. Animal population
densities are modeled as the proportion of fauna in the local eco-system
domesticated. An increase in
domestication is associated with a higher frequency of contact between host
populations (both human and animal).
No
modifications were made to the regional patch in the HDR model in order to
retain core-periphery differentiation[4] between
the complex local sedentary society (core) and simple regional sedentary
societies (periphery). The only
indicator of core-periphery difference is resource consumption and production
capacity since the regional system is modeled as an aggregated system of simple,
horticultural societies, which renders population density indicators invalid
(unless one assumes the number of societies in the regional system). Resource production and consumption is
greater in local societies with a moderate rate of domestication.
Inter-societal
relations, in the HDR model, only occur through emigration and warfare, thus, a
core assumption of the model is no positive relations. In context of the modified HDR model, the
demographic dynamics of the local society operates as a mediator in the
relationship between disease and inter-societal relations. According to the HDR model, the primary
factor driving warfare is the relative the degree of circumscription, which is
a function of regional population and resource consumption. The main effect of disease on
inter-societal relations is the dampening effecting on emigration, where
virulence operates as a check on demographic pressure. Further, domestication increasingly
improves consumption per capita, limiting the effect of internal conflict on
emigration. The expected relationship
between local disease dynamics, domestication, and warfare is a negative
association. The primary justification
for the association is centered on the demography of warfare and conflict in
early world-systems. Population
pressure over scarce resources results in higher frequencies of internal
conflict and emigration (Chase-Dunn and Hall 1997). However, in conditions with low population
pressure, due to the emergence of lethal pathogens, other regulations would
function as secondary mechanisms.
Therefore, in conditions with small populations distributed over
sufficient space, conflict is unlikely to arise at the local or systemic
level.
Three parameters are elucidated in
the analysis of the relationship between disease characteristics and
inter-societal relations: transmission risk, mortality, and domestication. Disease, migration, and warfare are
theorized as central mechanisms in the regulation of human populations in early
simple societies. In the transition to
more complex, agricultural societies, I argue that diseases becomes the primary
demographic regulator until the society develops endogenous and exogenous means
of handling disease (advanced health and medicine; sanitation; immunization;
etc.). Scenarios with disease as the
primary regulator, I predict little to no warfare or migration will occur,
which will result in little regional formation.
Modified HDR Model
The
methodological objective of this theoretical study is the development of a
mathematical formalization of the modified HDR model. Using the single society and world-system
versions of the HDR model (c.f. Apkarian 2010 et al), I added a generic SIR
modular.[5] Further,
an additional modification to the HDR model is the introduction of
domestication and animal population density.
Domestication is an exogenous rate with the range of 0 to 1, where 1
indicates that most, if not all, resources are locally cultivated in a specific
space. Domestication influences three
components of the modified HDR model.
First, domestication causes an upward shift in the local resource
reproduction rate. Second,
domestication increases the local harvesting rate of available resources. Lastly, domestication increases the animal
population density. The function for
animal population density is the following:
(3) ![]()
Where, a is
animal population density; r is the available local resources; l is the local
land size; and d is the domestication rate.
The
assumptions governing animal population density is that the eco-system is
composed half fauna and half flora.
Domestication operates as the percentage of local fauna that lives in
close proximity to human populations based on the spatial scale of the society
(land). According to these relations,
domestication plays a central role in reproducing human societies (increasing access
to greater resources and consumptions), while at the same time, creating the
conditions for the emergence of crowd disease (Diamond 1997). The
capture and domestication of wild animals facilitated the emergence of disease
pools where pathogens, adapted to animal host populations, were able to
transmit to densely populated human societies.
The
purpose of the formalized model is to explore the hypothetical relationship
between disease and inter-societal relations as demographic regulators. In order to test this relationship between
disease and inter-societal relations, I performed a series of simulations using
a formalized version of the HDR-disease model developed in STELLA 9.0.2[6]
(iseeSystems 2007). In total, ran 135
simulations based on 27 scenarios. Each
scenario was estimated 5 times in order to adjust for stochastic parameters in
the HDR model. Three parameters
defined the different scenarios: transmission risk, mortality, and
domestication. Each parameter was
assigned a low, medium, and high value.
For transmission risk, the low value was .02, the medium value was .5,
and the high value was 1. For
mortality, low value was .05, the medium value was .5, and then high value is
.9. Lastly, domestication ranged from
the low value, 0, to the high value 1.
The purpose of the different settings was to test the sensitivity of the
model to changes in pathogen strain.
Certain pathogens, such as influenza, are highly contagious and
transmission is high in dense populations, while other diseases, such as HIV,
have lower transmission risk, but higher mortality. Further, outside of pathogens, different
rate of domestication is used as an indicator of how variation in resource
production, as a result of human domestication, impacts the dynamics of disease
and inter-societal relations.
Model Dynamics: Disease and Inter-Societal Relations
Disease
The results of
the simulations confirmed the expected relationship between disease,
domestication, warfare, and migration.
In scenarios with medium domestication, where the proportion of resource
production exceeds 50% and local pathogens are highly virulent and transmission
risk is moderate to high, disease has a high risk of causing the local
population to go extinct. If
probability of mortality is less than 50%, the local population is regulated by
disease mortality. In conditions with
disease regulation, human populations experience a cyclical trend in emergence
and re-emergence of disease. Over the
course of 2000 iterations, the infection reproduction rate oscillates between
near-zero to about 100. Figure 3 (in
Appendix) shows the cyclical patterning of the number of infections per
iteration. A general interpretation of
the cyclical trending is the emergence of an epidemic, followed by a series of
endemic iterations, and re-emergence of epidemic. This type of cyclical dynamic is expected in
early populations without genetic adaptations for immunity to certain endemic
disease[7].
Warfare and Disease
A central
assumption in the original HDR model is that the regional area is depopulated
and can only be populated by migration from the local population. In order to test the model’s war dynamics, I
set the initial regional population to ten times that of the local
population. When the regional system is
populated, over successive iterations, warfare emerges as a secondary regulator
of local populations. Setting the
disease conditions to relatively medium and high conditions (transmission risk
= .7; mortality =.17) and moderate domestication (50% of resource production),
the emergence of disease and the cyclical trending of emergence and
re-emergence, is synchronized with the cycles of warfare. In iterations with the presence of disease
and warfare, warfare operates as a suppressant of the disease reproduction rate
(bringing R0 to <1 conditions for the iteration). Figure 4 (in the Appendix) shows the
cyclical relationship between warfare and disease reproduction rate under
favorable conditions for disease emergence.
In context of Figure 3, the counter-cyclical relationship appears to be
one of second-order regulation, where warfare, through death, operates
regulates the spread of disease. In iterations with warfare, the infection rate
lessens in magnitude, but maintains epidemic status. The interaction between these two dynamics
illustrates the growing complexity of demographic regulation in human societies,
however, warfare is assumed to be secondary to disease in regulating human
populations to sustainable levels.
Migration and Disease
A key dynamic in
the relationship between regional and local population is the migration of
local populations into regional space. According
to Hume et al (2003), early migration and spread of disease was a function of
overcrowding caused by sedentary lifestyles, urbanization, and
domestication. Under normal operating
conditions, the HDR model holds initial regional populations at zero and, over
successive iteration, the local population migrates into the regional system.
The
introduction of domestication into the local society patch changes the primary
drivers of migration in the society.
Low to medium domestication rates in the modified HDR model produces no
migration because the outward shift in resource production caused by
domestication diminishes the likelihood of internal conflict and raises
consumption per capita. As long as
diseases are present in the population, populations never exceed the critical
population density of two people per unit of land because disease death
operates as a regulator. In the
modified model, once the population exceeds this capacity, than migration increases. Once
migration occurs and the regional system populates, war dynamics tend to
co-determine populations with disease and other regulators.
Under the
same disease conditions simulated in the warfare tests, migration tests were
performed to observe the theoretical relationship between disease, migration,
and domestication. Scenarios with
high pathogen virulence (high transmission risk and low mortality) and little
to no domestication, diseases emerge very sporadically in the local population,
and migration occurs early in the simulation (prior to 500 years). In high virulence conditions with low to
moderate domestication, migration is unlikely to occur. Resource consumption in the local society
never results in the critical population density, thus, ecological needs are
met and overcrowding never occurs. Once
domestication rates exceed .75, migration occurs rapidly and frequent as a
result of overcrowding. Figure 5 (in
Appendix) shows the cyclical patterns in migration, warfare, and disease
reproduction. No real synchrony emerges
between the different regulators because each are functional and operative in
each iteration. In scenarios where all
three regulators (disease, migration, and warfare) are present, local and
regional populations has severe variation around a general equilibrium of about
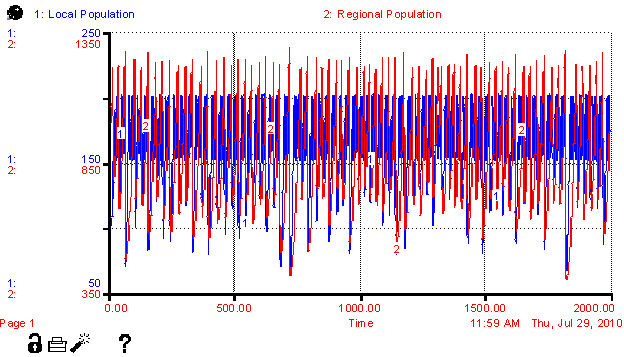
850 (regional) and 150 (local). Figure
6 (in Appendix) shows the severe oscillation of populations over 2000
iterations.
In
scenarios where the disease is assumed to have high to moderate mortality,
migration never occurs because disease death assumes the role of primary
demographic regulator. Besides
domestication, migration is highly dependent on the virulent mortality since
migration is a function of consumption, internal conflict, and population
density in the HDR model. If disease is
regulating the local population through high mortality, the regulatory function
of migration is unnecessary, thus, the world-system (local system plus regional
population) remains unpopulated. Only
in scenarios where the local population doesn’t experience high virulent
mortality, regional systems begin developing and disease becomes endemic or
follows a cyclical patterns of emergence and re-emergence.
Unlike warfare, migration in the
model does not follow a systemic pattern with disease dynamics. Rather, in scenarios with high population
density from domestication and moderate to high virulence, migration happens at
a higher frequency than disease dynamics and warfare. In this scenario, migration becomes the
primary demographic regulator and disease and warfare become secondary.
Discussion and Conclusion
The purpose of extending on previous mathematical
formalization of human social evolution is to introduce disease into the
theorization of long-term systemic evolution.
The role of disease in early human society, and with the advent of
domestication, has been central in regulating the relationship between human
populations and their eco-system. At
the nexus of humans-ecology, pathogens operate as environmental responses to
human action and represent a biological systems being co-determined by the
interaction between society and the environment. Using simulation techniques and mathematical
formalizations, I developed a simple addition to the HDR model aimed at testing
the human, system, and disease dynamics in scenarios of different pathogen
conditions.
The main emphasis has been on the
relationship between disease and inter-societal relations, specifically,
warfare and migration. The outcome of
the model suggests that disease, in early sedentary societies, functioned as a
secondary and primary regulator of human populations. Beyond resource consumption, the roles for
migration, inter-societal conflict, and disease are centered on maintaining an
equilibrium between human populations and local resources. Previous theorization (Apkarian et al 2010)
using the HDR model, found that endogenous (resource consumption) and exogenous
processes (war and migration) operated to produced equilibrium solutions with
variation. Under certain conditions,
the domestication and pathogen virulence created similar outcomes, but
variation and equilibrium was dictated by disease dynamics or disease killed
the populations.
The next step in the modeling process is
to include endogenous processes of pathogen adaptation to human host
populations. Over the course of
co-dependent evolution and human social evolution, pathogens have becoming
increasingly apt at responding to changes in their ecosystems, i.e. human
populations. Further, the emergence of
immunization, as both an adaptation of disease to human and human to disease
needs to be included in order to understand how pathogens have maintained an
endemic status in human societies.
Lastly, climate variation needs to be included in order to account for
the early emergence of temperature-dependent microorganism.
To account for the inter-societal
dimension of disease, the next model will include the process of diffusion
between two societies, where migrants of the local population are able to
spread diseases into surrounding populations.
The inter-societal transmission of disease is an important factor in
understanding the next major epidemic transition: pandemics. The intensification of inter-societal
connections in the modern world-system can be traced to earlier mergers of
disease pools in
References
Alizon,S.,
A. Hurford, and M. Van Baalen.
2009. “Virulence Evolution and
the Trade-Off
Hypothesis: History,
Apkarian,
Jacob, Jesse Fletcher, Robert A. Hanneman, Hiroko Inoue, Kirk Lawrence and
Christopher Chase-Dunn. 2010. “Human Demographic Regulators in Small-Scale
World-Systems.” IROWS Working Paper 41.
Armelagos,
George, A.H. Goodman, and K. Jacobs.
1991. “The Origins of
Agriculture:
Population Growth during a Period of
Declining Health.” Population and
Environment 13(l):9-22.
Armelagos,
George and Kathleen Barnes. 1999. “The Evolution of Human Disease and the Rise
of Allergy: Epidemiological Transitions.” Medical
Anthropology 18: 187-213.
Chase-Dunn,
Christopher Thomas Hall. 1997. Rise
and Demise: Comparing World-Systems.
Cockburn,
T.A. 1971. “Infectious Disease in
Ancient Populations.” Current
Anthropology
12(1):45-62.
Cohen,
Mark. 1977. The Food Crisis in
Prehistory: Overpopulation and the Origins of
Agriculture.
Crabtree,
Pam. 1993. “Early Animal Domestication in the Middle
East and
Archaeological
Theory and Method 5:
201-245.
de
Zulueta, J. 1994. “Malaria and
Ecosystems: From Prehistory to Posteradication.”
Parassitologia
36: 7–15.
Diamond,
Jared. 1997. Guns,
Germs, and Steel: The Fates of Human Societies.
W. Norton & Company.
_______. 2002.
“Evolution, Consequences and Future of Plant and Animal Domestication.”
Nature 418: 700-707.
Dobson,
Andrew and E. Robin Carper. 1996. “Infectious Diseases and Human Population
History.” Bioscience
46(2): 115-125.
Flannery,
K. 1969. “Origins and the Ecological Effects
of Early Domestication in
Near East.” Pp. 73-100 in The Domestication and Exploitation of Plants and Animals, eds.
Ucko, P. and G.W. Dimbleby.
Greger,
Michael. 2007. “The Human/Animal Interface: Emergence and
Resurgence of
Zoonotic Infectious Diseases.” Critical
Reviews in Microbiology 33: 243-299.
Gupta,
Anil. 2004. “Origin of Agriculture and Domestication of
Plants and Animals Linked to
Early Holocene Climate
Amelioration.” Current Science 87(1): 54-59.
Hannon,
Bruce and Matthias Ruth. 2009. "Basic Epidemic Models." In Dynamic Modeling of
Diseases and Pests.
Hume,
Jennifer, Emily Lyon, and Karen Day.
2003. “Human Migration,
Mosquitoes and the
Evolution of Plasmodium Falciparum.” Trends
in Parasitology 19(3): 144-149.
isee
Systems. 2007. STELLA 9.0.2.
Kermack,
W.O. and A.G. McKendrick. 1927. "A Contribution to the Mathematical
Theory of
Epidemics." Proceedings of the Royal Society of
of a Mathematical and Physical Character,
115 (772): 700-721.
McMichael,
A.J. 2004. “Environmental and Social Influences on
Emerging Infectious Diseases:
Past, Present and Future.” Philosophical
Transactions of the Royal Society B 359: 1049-1058.
McNeil,
William. 1976. Plagues
and Peoples.
Odum,
Eugene. 1975. Ecology: The Link Between the Natural and Social Sciences, 2nd
Edition.
Perkins,
Drexter. 1969. “Fauna of Catal Huyuk: Evidence for Early Cattle
Domestication in
Williams,
Michael. 2003. Deforesting the Earth:
From Prehistory to Global Crisis.
Wolfe,
Nathan, Claire Dunavan, and Jared Diamond.
2007. “Origins of Major Human
Infectious Diseases.” Nature
447: 297-283.
Woolhouse
Mark, Daniel Haydon, and Rustom Antia.
2005. “Emerging Pathogens: the
Epidemiology and Evolution of Species
Jumps.” Trends in Ecology and Evolution 20(5): 238-244.
Woolhouse,
Mark and Sonya Gowtage-Sequeria.
2005. “
Appendix
Figure 1. Iteration Model with Epidemics
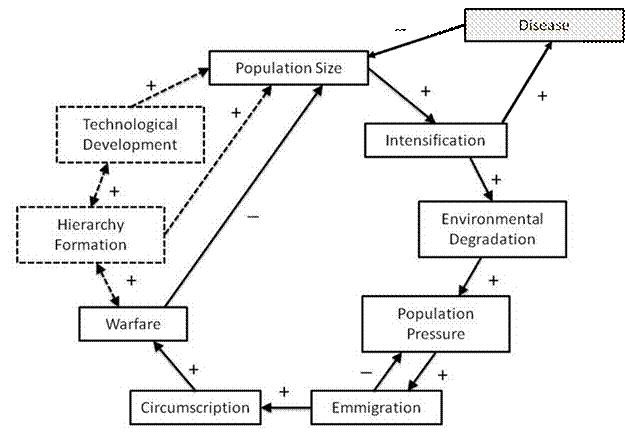
Figure 2. SIR Modular

Figure 3. Cyclical Epidemic-Endemic Trends in Local
Population
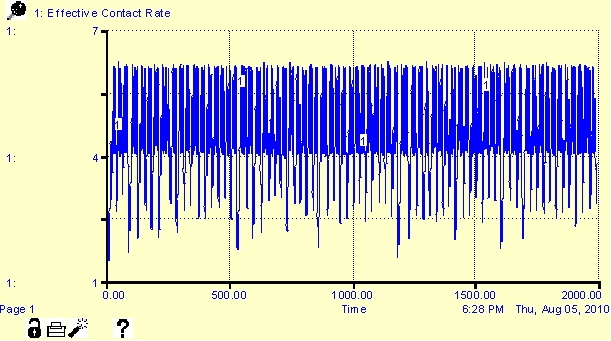
Figure 4. Synchrony of Warfare and Infection Rate
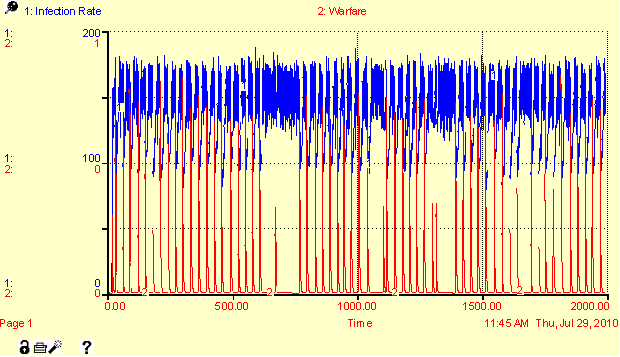
Figure 5. Demographic Regulators
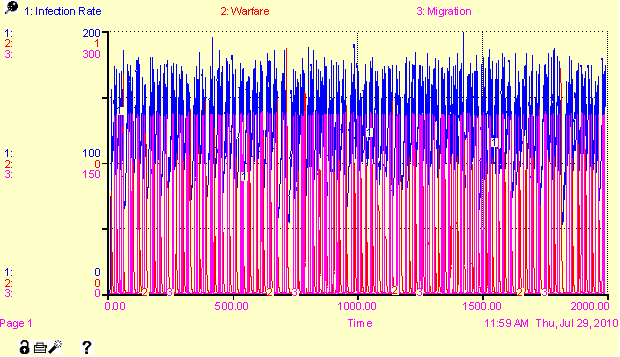
Figure 6. Regional and Local Population